
Príčiny selektívnej citlivosti plodín na podmienky pestovania a predovšetkým na rozdielnu koncentráciu vodíkových iónov vyjadrenú hodnotou pH pôdneho roztoku sú viacúrovňové. U pestovateľov pretrvávajú predstavy o rozdielnej citlivosti jednotlivých plodín na pH pôdy, čo sa aj uplatňuje pri pestovaní plodín. Odpoveď na otázku, prečo rastliny zdanlivo uprednostňujú určitý rozsah pH, nie je jednoduchá a vyžaduje jednotlivo analyzovať nasledovné parciálne otázky:
1/ Charakteristika pH z aspektu rozdielnej citlivosti rastlín
Pôdna reakcia je významným ukazovateľom vlastnosti pôdy, ktorá vplýva priamo a sprostredkovane na rast a vývin rastlín predovšetkým tým, že sa podieľa na mobilizácii živín v pôde, podmieňuje prijateľnosť živín z pôdy i hnojiva a určuje aj efektívnosť hnojenia plodín. Vyjadruje nasýtenosť sorpčného pôdneho komplexu a je výsledkom vzájomnej interakcie medzi pôdou, koreňovou mykorízou, aplikovanými hnojivami a koreňovým systémom plodín.
Jednotkou pôdnej reakcie je pH (pondus hydrogenii), ktorá predstavuje záporný dekadický logaritmus koncentrácie vodíkových iónov definovaný vzťahom:
pH = – log [ H+ ]
Pri pH = 7 koncentrácia H+ sa rovná 10-7 mol.dm-3. Pritom roztoky, ktoré pri 25 0C vykazujú hodnotu pH 7, patria medzi roztoky neutrálne a roztoky s pH pod 7 sú kyslé a nad pH 7 zásadité (alkalické). Iónový súčin vody vo forme koncentrácií H+ a OH– je vždy 10-14 mol2.dm-6.
Pôdna reakcia pôdy je závislá od prítomnosti v pôde: kyselín, zásad, obsahu výmenných katiónov, druhového zloženia a aktivity mikroflóry a intenzity mineralizácie organických látok, obsahu prijateľných živín z pôdnych zásob a hnojiva, ale aj in putu látok z atmosféry. Kyslosť pôdneho roztoku je významne podmienená predovšetkým prítomnosťou vodíkového iónu v pôdnom roztoku, ale aj obsahom výmenného hliníka Al-3 .
2/ Špecifické požiadavky plodín na pH
Vo všeobecnosti sa vychádza z predstavy, že rastliny sú prispôsobené na široký rozsah pH pôdy od 4 do 8. Avšak pre väčšinu plodín optimálne priemerne hodnoty sú v rozsahu od 5,5 do 7,5 . Manifestovaná toxicita je evidentná iba pri pH < 3 a nad pH 10, ktorá spôsobuje patologickú zmenu štruktúry koreňového systému. Rastliny vykazujú väčšiu odolnosť voči vysokým hodnotám pH. Existujú evidentné špecifiká plodín na optimálne hodnoty pH. Rozdiely medzi plodinami nie sú významné, spravidla sa líšia o jednu, eventuálne dve jednotky pH. Napríklad pri pestovaní zemiakov, ovsa, raže, ľanu a slnečnice hodnoty pH by nemali klesnúť pod 5,0. Naproti tomu pre pšenicu, jačmeň, kukuricu, fazuľu je žiaduce pH nad 6,0. Pre vičenec, kapustu, hrach a uhorky sa vyžadujú pH vyššie ako 6,5. Orientačné požiadavky jednotlivých plodín na pôdnu reakciu sú uvedené v Tab. 1. Opätovne treba zdôrazniť, že pH na rast plodín vplýva iba nepriamo – sprostredkovane cez deficit alebo nadbytok pre rastliny prístupných živín v pôdnom roztoku.
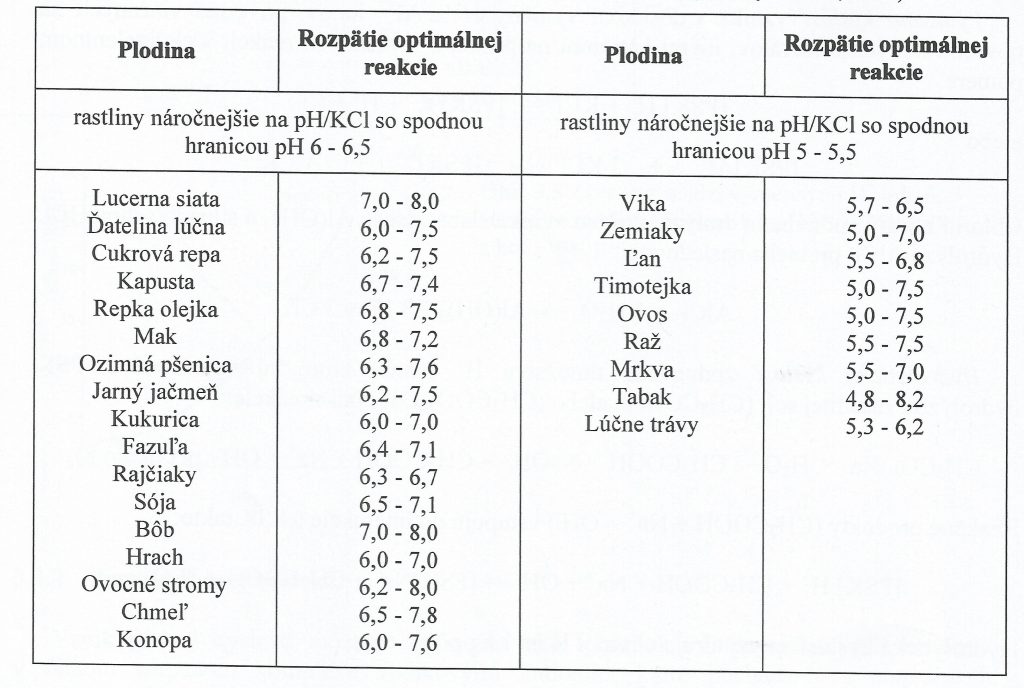
Tab. 1: Zdanlivé požiadavky plodín na pôdnu reakciu (podľa Masaryka a kol.,198O)
Pôdna reakcia významne vplýva na druhové zloženia a aktivitu pôdnej mikroflóry. Vo všeobecnosti sa predpokladá, že baktérie preferujú neutrálnu a mierne zásaditú reakciu a naproti tomu mikroskopické huby kyslú reakciu. Preto zmena pH pôdy vplýva na druhovú skladbu pôdnej mikroflóry, jej aktivitu, čo sa prejavuje v biologickej sorpcii živín a mineralizácii organických látok. Koreňová mykoríza vykazuje evidentne selektívne špecifikáa k jednotlivých druhom plodín. Koncentrácia vodíkových eventuálne hydroxylových iónov na rastové procesy poľných rastlín má širokospektrálny účinok, ktorý je možné vyjadriť účinnosťou procesov mobilizácie živín v pôde a príjmu porastom.
3/ Významná úloha pH pri realizácii procesov príjmu živín rastlinami
Mimoriadny význam pH pripadá na procesy príjmu živín. Prístupnosť živín a ich akumulácia v rastlinách je v značnej miere podmienená hodnotou pH pôdneho roztoku. Koncentrácia vodíkových a eventuálne hydroxylových iónov významne ovplyvňuje fyzikálno-chemické procesy sorpcie a desorpcie iónov, čo určuje mobilitu živín v pôdnom prostredí a vytvára priaznivé podmienky pre ich príjem.
Vychádza sa z predstavy, že hodnota pH vplýva na mikrobiálnu aktivitu, procesy mineralizácie organickej hmoty, rozpustnosť CO2, pričom zvýšená koncentrácia oxidu uhlíka nad 2 – 3 % v pôdnom roztoku znižuje príjem iónov v nasledovnom poradí : K+ > NH4+ > Fe3+ > Ca2+ > Mg2+. Pri vysokej hodnote pH dochádza ku intenzívnej sorpcii železa a viacerých mikroživín, čo spôsobuje pokles ich rozpustnosti a prístupnosti pre rastliny. Optimálny príjem fosforu je v oblasti pH 6,5 – 7,5. Naproti tomu v podmienkach pH 9 dochádza ku prerušeniu príjmu fosforu. Pre draslík sa vyžaduje pH nad 6,0 a pre vápnik a horčík nad 6,6. Optimálne hodnoty pre príjem dusíka sú v rozsahu pH 6 – 8, pričom pre príjem NO3– je < 7 pH a pre NH3 > 7.
Mikroživiny v pôde sú imobilizované vo forme nerozpustných zlúčenín, preto ich solubilita (rozpustnosť) a prienik do pôdneho roztoku je závislá predovšetkým od hodnoty pH. Vo všeobecnosti platí uzancia, že v kyslom prostredí stúpa rozpustnosť a tým aj prijateľnosť Fe, Mn, Zn, Cu, a B a naopak znižuje sa rozpustnosť a prístupnosť molybdénu pre rastliny.
Evidentné rozdiely v citlivosti rastlín sú iba zdanlivo na reakciu pôdneho roztoku, v skutočnosti je príčinou efekt deficitu alebo prebytku prístupných živín. Toto tvrdenie môžeme demonštrovať na príklade vysokej koncentrácie vodíkových iónov (veľmi nízke hodnoty pH), ktoré sú spravidla príčinou deficitu vápnika a čiastočne i fosforu a naproti tomu vysokého obsahu prístupného hliníku a mangánu v pôdnom roztoku. Z uvedeného je evidentné, že bezprostrednou príčinou toxicity plodín nie je vysoká koncentrácia H+, ale citlivosť plodín na deficit vápnika a toxicitu vysokej koncentrácie Al a Mn. Nadmerné koncentrácie hliníka v koreňoch vykazujú mimoriadne negatívny efekt na transport fosforu z koreňov do nadzemných orgánov, čo spôsobuje manifestovaný deficit fosforu. Reakcia plodín na deficit prístupného vápnika a nadmernú koncentráciu Al a Mn medzi druhmi plodín je veľmi rozdielna, čo môžeme demonštrovať na príklade dvoch botanicky odlišných plodín, akými je cukrová repa a zemiaky. V modelových podmienkach bolo dokázané, že cukrová repa a zemiaky rovnako vykazujú zvýšené nároky na obsah vápnika, avšak repa je relatívne odolná na vysoké koncentrácie hliníka. Naproti tomu porast zemiaka je citlivý na Mn a odolný na toxicitu spôsobenú hliníkom. Existuje analógia medzi zemiakom a druhmi plodín z čeľade krížokvetých. Jačmeň z hľadiska reakcie na Ca, Al a Mn čiastočne kopíruje vlastnosti repy cukrovej. Naproti tomu ovos v podmienkach nízkych hodnôt pH nevykazuje deficit vápnika (požiadavky na Ca sú nízke) a pritom je mimoriadne odolný na vysoké koncentrácie Al a Mn. Pravdepodobne preto existuje v praxi zaužívaná predstava o odolnosti porastu ovsa na nízke pH. Avšak skutočná podstata toho javu je v znížených nárokoch na vápnik a v odolnosti ovsa na vysoké koncentrácie Al a Mn.
Vo všeobecnosti sa predpokladá, že pre pestovanie lucerny, jačmeňa a repy cukrovej sú vhodné neutrálne a mierne kyslé pôdy (pH). Naproti tomu pšenica, hrach, bôb, vika a ďatelina plazivá sú schopné zabezpečiť dobru úrodu ako na neutrálnych, tak aj na kyslých pôdach. Je pozoruhodné, že plodiny ako ďatelina biela, ovos, raž, lupina a zemiaky dobre znášajú pôdy veľmi kyslé, ale aj neutrálne. Z uvedeného je evidentné, že tolerancia plodín na pH je pomerne široká. Aplikácia maštaľného hnoja zmierňuje negatívny účinok nízkych hodnôt pH. Pozoruje sa závislosť medzi zvýšenou odolnosťou na nízke hodnoty pH v podmienkach chladného a vlhkého počasia a naopak teplé počasie a vlahový deficit zvyšuje negatívny dopad zvýšených hodnôt pH. Špecifická reakcia tráv na pH sa prejavuje v ekosystémov trvalých trávnych porastov, kde dochádza ku vzájomnej konkurencii medzi druhmi plodín o prístupné zdroje živín. Predpokladáme, že príčina vplyvu vysokých hodnôt pH (alkalická pôdna reakcia) na retardáciu rastu porastu taktiež nie je spôsobená zvýšenou koncentráciou hydroxylových iónov, ale najmä deficitom Fe, Mn, B, P a pravdepodobne i ďalších živín.

Vo všeobecnosti sa vychádza z predstavy, že rastliny sú prispôsobené na široký rozsah pH pôdy od 4 do 8.
4/ Regulačná úloha pH v činnosti iónogénnych púmp
I napriek uvedenému treba konštatovať, že regulačná funkcia pH sa prejavuje nie iba v tvorbe prístupných zdrojov živín, ale aj procesoch mechanizmu príjmu a akumulácie živín v rastlinách.
Pochopenia regulačnej funkcie pH v príjme živín je možné demonštrovať na príklade aktívneho príjmu dusičnanov a amoniaku proti koncentračnému gradientu, ktorý sa uskutočňuje činnosťou iónogénnej pumpy. Výsledky znázornené na obr. 1 demonštrujú rozdielny účinok pH na príjem chemických foriem dusíka.
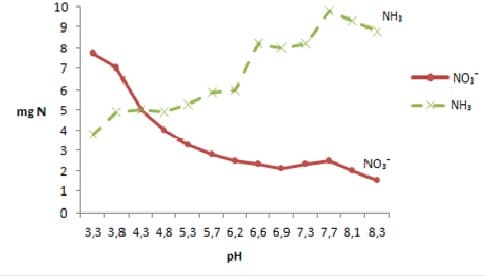
Obr. 1: Vplyv pH na dynamiku príjmu dusičnanov a amoniaku.
V závislosti od pH sa striedavo nastavuje pracovný režim „pumpy“ na „prečerpávanie“ NO3– alebo NH4+. Podľa predstáv Dejeagerrera (1981) sa na 61 prijatých katiónov (NH4+) v antiporte s vodíkovým iónom do prostredia uvoľní 85 H+ , ktoré spôsobuje zníženie pH pôdneho roztoku (< 7,0 pH), čo sa následne prejaví v stimulácii príjmu (influxu) dusičnanov v antiporte s OH–. Akumulácia dusičnanov a následná redukcia na amoniak v procese jeho asimilácie vytvára potenciálne zdroje hydroxylových iónov, ktoré podmieňujú nárast pH (alkalické prostredie) pôdneho roztoku ( > 7,0 pH), čím sa vytvárajú priaznivé podmienky na prednostný príjem amoniaku.
Uvedený dômyselný mechanizmus striedavého príjmu katiónov a aniónov na príklade iónov dusíka, ktorý je regulovaný pH, umožňuje stálu hodnotu pH cytoplazmy na úrovni 7,0. Pritom tento mechanizmus zároveň prispieva ku eliminácii toxicity amoniaku tým, že ho „prečerpáva“ z cytoplazmy do pôdneho roztoku, čo je evidentné z výsledkov znázornených na obr. 2.
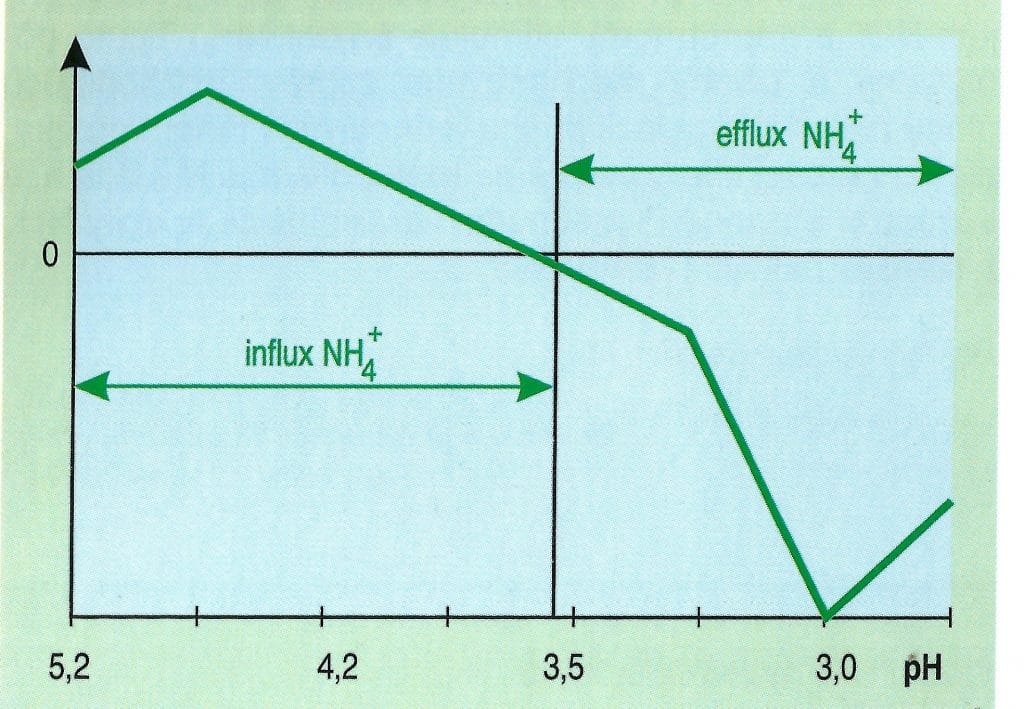
Obr. 2: Vplyv pH na aktívny influx a efflux.
Analyzovaný model príjmu chemických foriem dusíka je dôkazom regulačnej funkcie pH na realizácii akumulácie živín. Aktívny mechanizmus príjmu živín vyžaduje spotrebu metabolickej energie a činnosť katalytických systémov, ktorých účinnosť a kapacita pri jednotlivých plodinách je rozdielna.
Medzi plodinami sú evidentné odlišnosti v schopnosti buniek semena a klíčiacej rastliny odolávať amoniakálnej toxicite, eventuálne toxicite iných iónov, napríklad chlóru, hliníka, mangánu a ďalších.
Je zrejmé, že rastliny sa musia brániť proti vysokej koncentrácii iónov. Hmotná podstata odolnosti rastlín na zvýšenú koncentráciu amoniaku je daná predovšetkým obsahom v semene metabolizovateľných sacharidov, ktoré v procese klíčenia poskytujú prekurzory (organické oxo-kyseliny) nevyhnutné na imobilizáciu amoniaku (asimilácia amoniaku) vo forme aminokyselín. Pestovatelia dobre vedia o mimoriadnej citlivosti klíčkov repy cukrovej na prítomnosť amoniaku v pôde. I napriek tomu, že repa produkuje buľvy s vysokým obsahom sacharózy, semeno repy cukrovej, na rozdiel od semien iných plodín (najmä obilnín), je chudobné na obsah škrobu, čo je hlavnou a jedinou príčinou zvýšenej citlivosti repy na koncentráciu amoniaku. V dôsledku uvedeného pri pestovaní repy cukrovej je nežiaduce aplikovať amoniakálne hnojivá tesne pred výsevom alebo v čase sejby. I na tomto príklade vidíme analógiu v rozdielnej reakcii jednotlivých druhov plodín na podmienky prostredia, pričom odolnosť rastlín je výlučne determinovaná endogénnymi – geneticky podmienenými procesmi.
5/ Vplyv pH na aktivitu enzýmov a endogénne metabolické a biosyntetické procesy
Jednotlivé enzýmy vykazujú prísnu katalytickú aktivitu iba vo vymedzenom rozsahu pH, čo predstavuje špecifikum katalytických systémov. Väčšina enzýmov vykazuje katalytickú aktivitu v úzkom rozmedzí medzi 5 a 9 pH. Efekt pH vplýva selektívne na väzbu substrátu na enzým, čo je spôsobené disociáciou molekuly substrátu a zmenou štruktúry bielkoviny. Tým, že enzýmy sú bielkoviny, potom i katalytické procesy sú prísne geneticky determinované. Z uvedeného je evidentné, že pH sa podieľa na regulácii produkčnej výkonnosti porastu.
Mimoriadne špecifická funkcia koncentrácie protónov a najmä gradient iónov rozhoduje o účinnosti transformácie foriem energie v biologických systémoch. Tým, že živé organizmy vrátane rastlín patria medzi izotermické systémy, sú schopné transformovať iba energiu koncentračného gradientu, ktorý sa vytvára na rozhraní membrán mitochondrií a chloroplastov. Univerzálnym energetickým iónom je vodíkový ión – H+ (protón). Z uvedeného vyplýva, že rozdielna koncentrácia H+ na rozhraní nepriepustnej membrány generuje protónový gradient ΔH+, ktorý je ekvivalentom ΔpH a je hybnou silou transformácie oxidačnej (produkt katabolických reakcií – oxidačná fosforylácia) alebo fotosyntetickej energie (fotosyntetická fosforylácia). Premena energie sa uskutočňuje nadmolekulárnym katalytickým systémom adenozin trifosfatázy (H+– ATP- áza) a produktom transformácie energie protónového gradientu – ΔH+ je chemická forma energie (ATP). Bioenergetický systém adenozín trifosfatázy predstavuje biologický molekulový elektrochemický „stroj“, ktorý prečerpáva cez membránu protóny (protónová pumpa) a zároveň vykonáva prácu, ktorej výsledkom je univerzálny biologický energetický produkt ATP.
Záver
Na záver treba konštatovať, že všetky kultúrne plodiny vznikli výberom a neskôr i šľachtením divokých typov a foriem rastlín, ktoré rástli v konkrétnych prirodzených agroekologických podmienkach. Počas dlhodobého obdobia evolúcie sa rastliny adaptovali na špecifické prírodné živinové podmienky prostredia, umožnili sformovať adekvátne anabolické a katabolické systémy, ktoré najplnšie zabezpečujú potreby rastlín. Medzi významné pôdno-ekologické podmienky patrí teplota, vodný režim, mineralogické zloženie pôdneho substrátu, obsah organických látok, prístupných živín, skladba a aktivita koreňovej mykorízy, koncentrácia CO2 a O2 a pH pôdneho roztoku. Afinita rastlín na špecifické podmienky prostredia je determinovaná endogénnymi procesmi, ktoré vytvárajú optimálne parametre pre rast a vývin i formovanie biomasy. Pri hodnotení významu pH je potrebné diferencovať vplyv pôdnej reakcie na mobilizáciu živín v pôde a na regulačnú úlohu koncentrácie vodíkových iónov na endogénne procesy v rastlinách.
Medzi plodinami existujú čiastočné rozdiely v selektívnej citlivosti na deficit živín, ale aj na vysoké koncentrácie solí a osobitne Na, Cl, NH3, Al, Mn, NO3– , H+ a OH– . Odolnosť alebo adaptabilita jednotlivých plodín na podmienky prostredia je spojená s existenciou endogénnych obranných mechanizmov buniek koreňového systému a schopnosťou účinne vylučovať (efflux) nežiaduci prebytok iónov z cytoplazmy do pôdneho roztoku, alebo ich imobilizovať vo forme nerozpustných zlúčenín (organominerálnych komplexov). Metabolické a syntetické procesy v rastlinách patria medzi enzymatické systémy, ktoré sú prísne geneticky determinované.
Existujú evidentné rozdiely medzi plodinami v požiadavkách na optimálne prostredie ich pestovania, dané obsahom živín a rozsahom pH. Pričom špecifické agroekologicke podmienky rastu sú charakteristické pre určitý druh plodín. Medzi rastlinami a prostredím sú vytvorene vzájomné väzby. Rastliny z prostredia prijímajú vodu, CO2 a živiny a do prostredia vylučujú okrem minerálnych látok (OH–, H+, Na+, Cl–, K+, NO3–, NH4+) aj organické látky (monosacharidy, aminokyseliny, organické kyseliny), čím prispievajú ku tvorbe priaznivých živinových podmienok pre optimálny rast.
Vo všeobecnosti dominuje názor, že odlišná citlivosť rozdielnych druhov plodín na pH pôdneho prostredia sa prejavuje nepriamo tým, že pôdna reakcia vplýva na chemické a biologické procesy mobilizácie živín a ich prístupnosti pre rastliny. V dôsledku uvedeného konkrétne podmienky pH sú zodpovedné predovšetkým za koncentráciu iónov, najmä živín v pôdnom roztoku, ktorá je na úrovni optimálnej, deficitnej alebo toxickej.
Na základe analyzovaných skutočností je žiaduce korigovať názory na citlivosť, eventuálne odolnosť plodín na podmienky pH. V skutočnosti za prirodzených podmienok pestovania plodín (pH 4 – 8) zdanlivo odlišná citlivosť plodín nie je spôsobená koncentráciou H+ a OH–, ale predovšetkým deficitom alebo nadlimitnou koncentráciou jednotlivých živín. Plodiny vykazujú špecifickú selektívnu reaktivitu a tým i schopnosť adaptácie iba na prítomnosť konkrétnych živín.
Autor: prof. Ing. I. Michalík, DrSc.







