
Z hľadiska výživy kukurice siatej mikroelementmi je známe, že kukurica veľmi dobre reaguje na hnojenie meďou a zinkom. Dobre na hnojenie bórom, molybdénom, mangánom a železom. Hnojenie inými mikroelementmi je málo efektívne. Veľkosť aplikačnej dávky závisí od zásoby daného prvku v pôde, jeho potreby na jednotku úrody a výšky plánovanej úrody.
Aplikácia väčších dávok pevných Cu a Zn hnojív do pôdy by sa mala realizovať v šesťročných intervaloch pri jesennej príprave pôdy – 1/3 pri podmietke a 2/3 pri orbe. Menšie dávky sa aplikujú v trojročných intervaloch a to 2/3 pri jesennej a 1/3 pri jarnej príprave pôdy. Malé dávky (v pevných i kvapalných hnojivách) sa aplikujú pri jarnej príprave pôdy.
Kvapalné hnojivá je efektívne používať najmä pred sejbou a počas vegetácie kukurice. Hnojenie počas vegetácie sa realizuje na základe rozborov rastlín. Ich odber môže byť v rôznych rastových fázach a pri rôznych výškach porastov. Výsledky rozborov sa porovnajú s údajmi uvedenými v tabuľkách 3 a 4. V prípade zistenia veľmi nízkej, resp. nízkej zásoby niektorého mikroelementu, je nutné ho dodať. Treba mať na zreteli, že čím neskôr sa vykoná korekcia zlého výživného stavu, tým je efekt nápravy menší.
Aplikačné dávky zinku pri foliárnej výžive sú najčastejšie na úrovni 20 až 500 g.ha-1, pričom sa Zn aplikuje zvyčajne dva až trikrát počas vegetácie. Existuje viacero poznatkov s postrekovaním rastlín dávkou 2 – 5 kg.ha-1 heptahydrátu síranu zinočnatého (23,5 až 35 % obsah Zn) vo forme 1 % roztoku, čím sa dodá 500 až 1 kg čistého zinku. Pri výraznom deficite zinku v rastlinách ho niektorí farmári aplikujú foliárne v celkovej dávke prevyšujúcej 1 kg.ha-1. Celková aplikačná dávka Zn pri foliárnej výžive nezvykne v podmienkach Slovenska presiahnuť 1,5 kg.ha-1.
Množstvo zinku prijatého rastlinami závisí od druhu pestovaných rastlín. Priemerný obsah Zn v poľných plodinách sa pohybuje od 20 do 100 mg.kg-1. Obsah pod 20 mg.kg-1 Zn na kilogram sušiny rastliny sa považuje za indikujúci nedostatok zinku a obsah nad 200 mg.kg-1 Zn sa považuje za toxický.
Význam zinku pre rastlinu je viacnásobný. Podporuje syntézu tryptofánu, ktorý je východiskovou látkou pre tvorbu auxínov. Je nevyhnutý z pohľadu integrity bunkových membrán. Pozitívne vplýva na intenzitu fotosyntézy, zlepšuje rast letorastov a zintenzívňuje príjem N, P a K, následkom čoho zvyšuje úrodu a zlepšuje jej kvalitu. V rastlinách stúpa obsah kyseliny L-askorbovej, ale aj iných organických kyselín, ako napríklad kyseliny vinnej a jablčnej v plodoch ovocia.
Napriek existencii viacerých poznatkov o vplyve nedostatku zinku, ale aj iných živín na tvorbu fytomasy kukurice siatej, sa na SPU v Nitre založil nádobový pokus, v ktorom sa sledoval vplyv deficitu 9 živín (5 makroelementov a 4 mikroelementov) na tvorbu fytomasy kukurice siatej, s cieľom presne kvantifikovať vplyv nedostatku jednotlivých prvkov na tvorbu slamy a šúľkov spolu so zrnom.
Pokus bol založený 13. mája. Do nádob tvaru valca vysokých 52 cm s priemerom 38 cm sa navážilo 13 kg zmesi pozostávajúcej z 3 kilogramov perlitu a 10 kg piesku. Agrochemické parametre použitých materiálov uvádza tabuľka 1. Takto pripravené nádoby sa zaliali destilovanou vodou na úroveň 40 % PVK. Následne sa zaliali Knopovými roztokmi do úrovne 70 % PVK. Knopov roztok obsahoval tieto živiny: N, P, K, Ca, Mg, S, Mn, Zn, Cu, Mo, Fe, B, Na, Cl. Celkovo bolo pripravených 10 variantov, 10 roztokov: Ú = úplný živný roztok, – N = živný roztok bez dusíka, – P = živný roztok bez fosforu, – K = živný roztok bez draslíka, – Mg = živný roztok bez horčíka, – S = živný roztok bez síry, – Cu = živný roztok bez medi, – Zn = živný roztok bez zinku, – Fe = živný roztok bez železa, – Mn = živný roztok bez mangánu. Pokus bol založený metódou náhodného usporiadania nádob s päťnásobným opakovaním. Keďže nádoby boli umiestnené vo vegetačnej klietke, hrozilo, že pravidelné, denné zalievanie nádob živnými roztokmi by pri veľkom výpare vody spôsobilo výrazné zasolenie substrátu. Z toho dôvodu sa pripravili presné objemy živných roztokov podľa Knopa, ktorými sa do polovice júna dodali živiny na vytvorenie 6 tonovej úrody zrna kukurice siatej pri kalkulácii 60 % využiteľnosti dusíka. Následne sa 4 týždne zalievalo len destilovanou vodou. V polovici júla bolo zrejmé, že absolútny deficit najmä makroelementov, ale aj mikroelementov spôsobí predčasné ukončenie vegetácie daných rastlín. Z toho dôvodu sa za účelom predĺženia života rastlín od 20. júla do všetkých nádob raz za týždeň (celkovo 8 krát) pridával úplný Knopov roztok o objeme 500 ml, čo umožnilo ukončiť pokus až v septembri. Na konci pokusu sa vyhodnotila tvorba nadzemnej fytomasy, hmotnosť slamy a hmotnosť zrna spolu so šúľkom.

Zinok podporuje syntézu tryptofánu, ktorý je východiskovou látkou pre tvorbu auxínov.
Z piatich sledovaných makroelementov najväčší vplyv na tvorbu celej nadzemnej fytomasy mal dusík. Jeho nedostatok znížil tvorbu nadzemnej fytomasy o 82,89 %, čo vysvetľuje prečo farmári i pri nedostatku financií na hnojivá, si takmer vždy nájdu finančné zdroje na nákup dusíkatých hnojív (obr. 1). Z obrázkov 2 a 3 vyplýva, že nedostatok dusíka viac ovplyvnil tvorbu generatívnych orgánov ako vegetatívnych. Nedostatok N rezultoval v nevytvorení samičích pohlavných orgánov, t. j. nevytvorili sa šúľky, čím sa nemohli vytvoriť ani zrná. Hmotnosť zŕn bola o 100,00 % a hmotnosť slamy o 65,25 % menšia v porovnaní s kontrolným variantom, na ktorom sa realizovala úplná výživa.
Nedostatok jednotlivých makroelementov sa na tvorbe generatívnych a vegetatívnych orgánov kukurice siatej neprejavil rovnako. Najmenší vplyv sa zaznamenal pri síre. Nedostatok síry znížil tvorbu nadzemnej fytomasy kukurice siatej len o 15,81 % a hmotnosť šúľka spolu so zrnom znížil o 22,57 %. Zistené korešponduje s poznatkom, že ak rastliny rastú v podmienkach nedostatku síry v pôde a dostatku síry vo vzduchu (nie nadbytku), dokážu prostredníctvom listov prijať až 90 % z jej celkového množstva v rastline. Poradie vplyvu nedostatku sledovaných makroelementov na tvorbu generatívnych i vegetatívnych orgánov kukurice siatej (Graf 1, 2 a 3) bolo nasledovné N > P > K > Mg > S, pričom deficit sa výraznejšie prejavil na tvorbe generatívnych orgánov ako na tvorbe slamy.
-
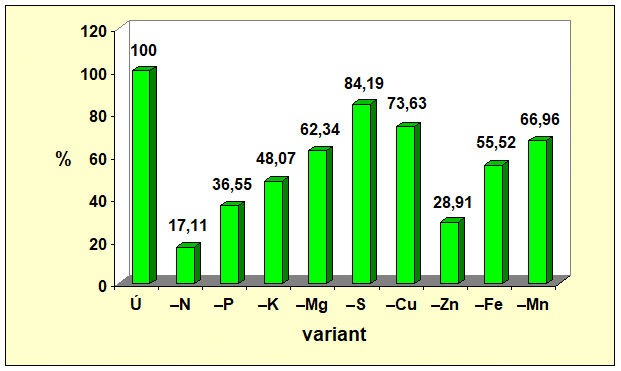
- Graf 1: Vplyv úplnej a deficitnej výživy vybranými makro- a mikroelementmi na hmotnosť nadzemnej fytomasy kukurice siatej.
-
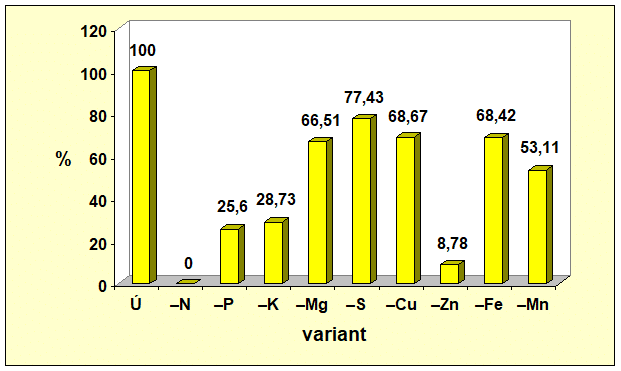
- Graf 2: Vplyv úplnej a deficitnej výživy vybranými makro- a mikroelementmi na hmotnosť šúľkov spolu so zrnom kukurice siatej.
-
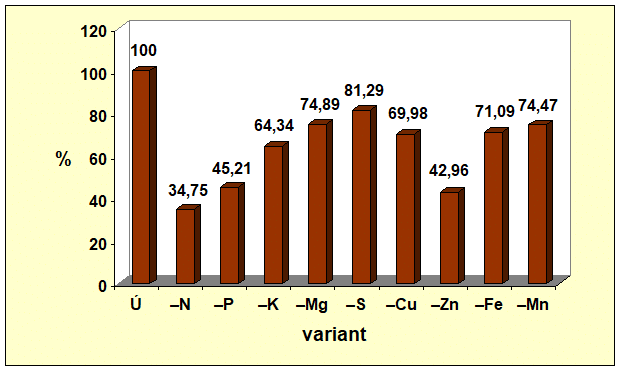
- Graf 3: Vplyv úplnej a deficitnej výživy vybranými makro- a mikroelementmi na hmotnosť slamy kukurice siatej.
Údaje v grafoch 1 až 3 vypovedajú, že zo štyroch testovaných mikroelementov, najväčší vplyv na tvorbu generatívnych i vegetatívnych orgánov mal zinok. Deficit zinku sa na tvorbe úrody zŕn podieľal významnejšie (91,22 %) ako na tvorbe slamy (57,04 %). Vplyv nedostatku Zn na tvorbu zŕn bol výraznejší ako vplyv fosforu, draslíka, horčíka či síry. Zistené poukazuje na významnú úlohu zinku pri dosahovaní vysokých úrod zrna kukurice siatej.
Zo všetkých 9 testovaných prvkov, úrodu zŕn kukurice siatej najviac negatívne ovplyvnil nedostatok N, potom nedostatok Zn a následne P, K, Mn, Mg, Fe, Cu, S. Tvorbu slamy taktiež najviac negatívne ovplyvnil nedostatok N, potom Zn, P, K, Cu, Fe, Mn, Mg, S.
Pri realizácii predloženého pokusu sa zaznamenal jeden menej známy poznatok: vo variante s deficitom draslíka v pestovateľskom substráte sa, na rozdiel od ostatných variantov, nedopestovalo žlté zrno, ale zrno žíhané, majúce červené až fialové prúžky, prípadne časť zrna bola zafarbená do sivočierna, resp. sfarbenie niektorých zŕn bolo tmavomodré (obr. 4). Pravdepodobnou príčinou objavenia sa červených až fialových prúžkov, tmavomodrej farby zrna bola skutočnosť, že draslík významne ovplyvňuje glycidový metabolizmus, pričom pri jeho poruchách sa v nadmernej miere tvoria antokyány. Antokyány sú vodou rozpustné pigmenty, ktorých farba môže byť červená, fialová, modrá.

Červeno až modrofialovo, resp. sivočierno žíhané zrná kukurice siatej a neopelená špička ako dôsledok nedostatku draslíka.
Okrem doposiaľ nám neznámeho zaznamenania žíhaného zafarbenia zŕn kukurice v dôsledku nedostatku draslíka sa zaznamenal i dlhodobo známy poznatok o nedoopelení špičky kukuričného šúľka (obr. 4) v dôsledku nedostatku draslíka.
Prezentované poznatky spresnili (číselne vyjadrili) význam racionálnej výživy kukurice siatej vybranými makroelementmi a mikroelementmi (N, P, K, Mg, S, Cu, Zn, Fe a Mn) a zároveň priniesli unikátny poznatok o zmene farby kukuričného zrna v dôsledku výrazného nedostatku draslíka v pestovateľskom médiu (v pôde).
Tab. 1: Agrochemické parametre použitého piesku a perlitu.
| Materiál | pH | Nan | P | K | Ca | Mg | S | Zn | Fe | Mn | Cu | Obj. hm. |
| mg.kg-1 | g/cm3 | |||||||||||
| Perlit | 7,95 | 10,3 | 5,0 | 175 | 300 | 113,3 | 5,0 | 0,69 | 10,98 | 0,38 | 0,32 | 0,18 |
| Piesok | 6,79 | 5,4 | 4,4 | 15 | 72 | 19,0 | 0,7 | 0,20 | 1,86 | 0,65 | 0,02 | 1,58 |
Tab. 2: Odber Zn a Cu úrodou jednej tony zrna a príslušným množstvom vedľajšieho produktu kukurice siatej.
| Produkty a ich zvyčajné pomery | Cu | Zn |
| g.t-1 zrna | ||
| Zrno 1 | 4,29 | 28,89 |
| Kôrovie 1,2 | 4,91 | 17,41 |
| Zrno + adekvátne množstvo kôrovia | 9,20 | 46,30 |
Zdroj: Neuberg et al., 1990
Tab. 3: Kritériá hodnotenia obsahu mikroelementov v kukurici siatej (Neuberg et al., 1990).
|
Prvok |
Rastová fáza
(doba odberu) |
Analyzovaná
časť |
Hodnotenie | ||||
| VN* | N | S | V | VV | |||
| mg.kg-1sušiny | |||||||
| B |
pri výške 20 – 30 cm |
nadzemná hmota |
2,0 – 5,0 | 5 – 25 | >25 | ||
| Cu | <5 | 5 – 20 | >20 | ||||
| Mn | < 30 | 30 – 300 | >300 | ||||
| Mo | <0,3 | 0,3 – 1,0 | >1,0 | ||||
| Zn | <16 | 16 – 23 | 23 – 56 | >56 | |||
*VN – veľmi nízky, N – nízky, S – stredný, V – vysoký, VV – veľmi vysoký
Tab. 4: Adekvátny obsah mikroelementov v rastlinách kukurice siatej (Fageria et al., 1991) .
| Prvok |
Rastové štádium |
Časť rastliny |
Primeraný
obsah (mg.kg-1) |
|
Zn |
30 – 40 dní po vzídení (klíčení) | celá nadzemná fytomasa | 20 – 50 |
| pred metaním (tvorbou metliny) | listová čepeľ pod praslenom
(zatočená dolná 1/3 posledného listu |
15 – 60 | |
| tvorba blizien samičích kvetov (fúzov) | šúľková listová čepeľ | 20 – 70 | |
|
Cu |
30 – 40 dní po vzídení (klíčení) | celá nadzemná fytomasa | 7 – 20 |
| pred metaním (tvorbou metliny) | listová čepeľ pod praslenom
(zatočená dolná 1/3 posledného listu) |
3 – 15 | |
| tvorba blizien samičích kvetov (fúzov) | šúľková listová čepeľ | 6 – 20 | |
|
Mn |
30 – 40 dní po vzídení (klíčení) | celá nadzemná fytomasa | 50 – 160 |
| pred metaním (tvorbou metliny) | listová čepeľ pod praslenom
(zatočená dolná 1/3 posledného listu) |
20 – 300 | |
| tvorba blizien samičích kvetov (fúzov) | šúľková listová čepeľ | 20 – 150 | |
|
Fe |
30 – 40 dní po vzídení (klíčení) | celá nadzemná fytomasa | 50 – 300 |
| pred metaním (tvorbou metliny) | listová čepeľ pod praslenom
(zatočená dolná 1/3 posledného listu) |
30 – 200 | |
| tvorba blizien samičích kvetov (fúzov) | šúľková listová čepeľ | 21 – 250 | |
|
B |
30 – 40 dní po vzídení (klíčení) | celá nadzemná fytomasa | 7 – 25 |
| pred metaním (tvorbou metliny) | listová čepeľ pod praslenom
(zatočená dolná 1/3 posledného listu) |
4 – 25 | |
| tvorba blizien samičích kvetov (fúzov) | šúľková listová čepeľ | 6 – 20 | |
|
Mo |
30 – 40 dní po vzídení (klíčení) | celá nadzemná fytomasa | 0,1 – 10 |
| pred metaním (tvorbou metliny) | listová čepeľ pod praslenom
(zatočená dolná 1/3 posledného listu) |
0,1 – 3,0 | |
| tvorba blizien samičích kvetov (fúzov) | šúľková listová čepeľ | > 0,2 |
klasová = šúľková
Autor: prof. Ing. P. Kováčik, PhD., SPU v Nitre







